 Scansoriopterygidae (meaning “creeping wings”) is an extinct family of climbing and gliding maniraptoran dinosaurs . Scansoriopterygids are known from five well-preserved fossils, representing four species, found in the fossil beds of the Tiaojishan Formation (dating from the Middle to Late Jurassic Period ) of Liaoning and Hebei , China :
Scansoriopterygidae (meaning “creeping wings”) is an extinct family of climbing and gliding maniraptoran dinosaurs . Scansoriopterygids are known from five well-preserved fossils, representing four species, found in the fossil beds of the Tiaojishan Formation (dating from the Middle to Late Jurassic Period ) of Liaoning and Hebei , China :

Scansoriopteryx heilmanni (and its likely synonym Epidendrosaurus ninchengensis ) was the first non-avian dinosaur discovered that had clear adaptations to an arboreal or semi-arboreal lifestyle: it is likely that they spent much of their time in trees. Both specimens displayed characteristics that indicated they were juveniles, making it difficult to determine their exact relationship to other dinosaurs and non-avian birds . It was not until the description of Epidexipteryx hui in 2008 that an adult specimen was known. In 2015 the discovery of another larger adult specimen belonging to the Yi qi speciesshowed that scansoriopterygids were not only climbers but also had adaptations that could have been used for gliding flight. The recently discovered (in 2019) Ambopteryx also demonstrates this . The previously described Pedopenna may also be a member of this clade. [1]

Scansoriopterygians are among the smallest known non-avian dinosaurs. Juvenile specimens of Scansoriopteryx are about the size of house sparrows , [2] about 16 cm (6.3 in) long, while the adult type specimen of Epidexipteryx is about the size of a pigeon , about 25 cm (10 in) long. 9.8 inches) (excluding tail feathers). [3]
Scansoriopterygids differ from other theropod dinosaurs in part by their extremely long third toe, which was longer than the first and second digits on the hand. In all other known theropods, the second toe is the longest. At least two species, Yi and Ambopteryx , also had a long “styliform” bone growing from the wrist, which, together with the third finger, helped support a bat-like wing membrane used for gliding. This use of a long finger to support a wing membrane is only superficially similar to the wing arrangement in pterosaurs , although physically it is more bat-like. [4]
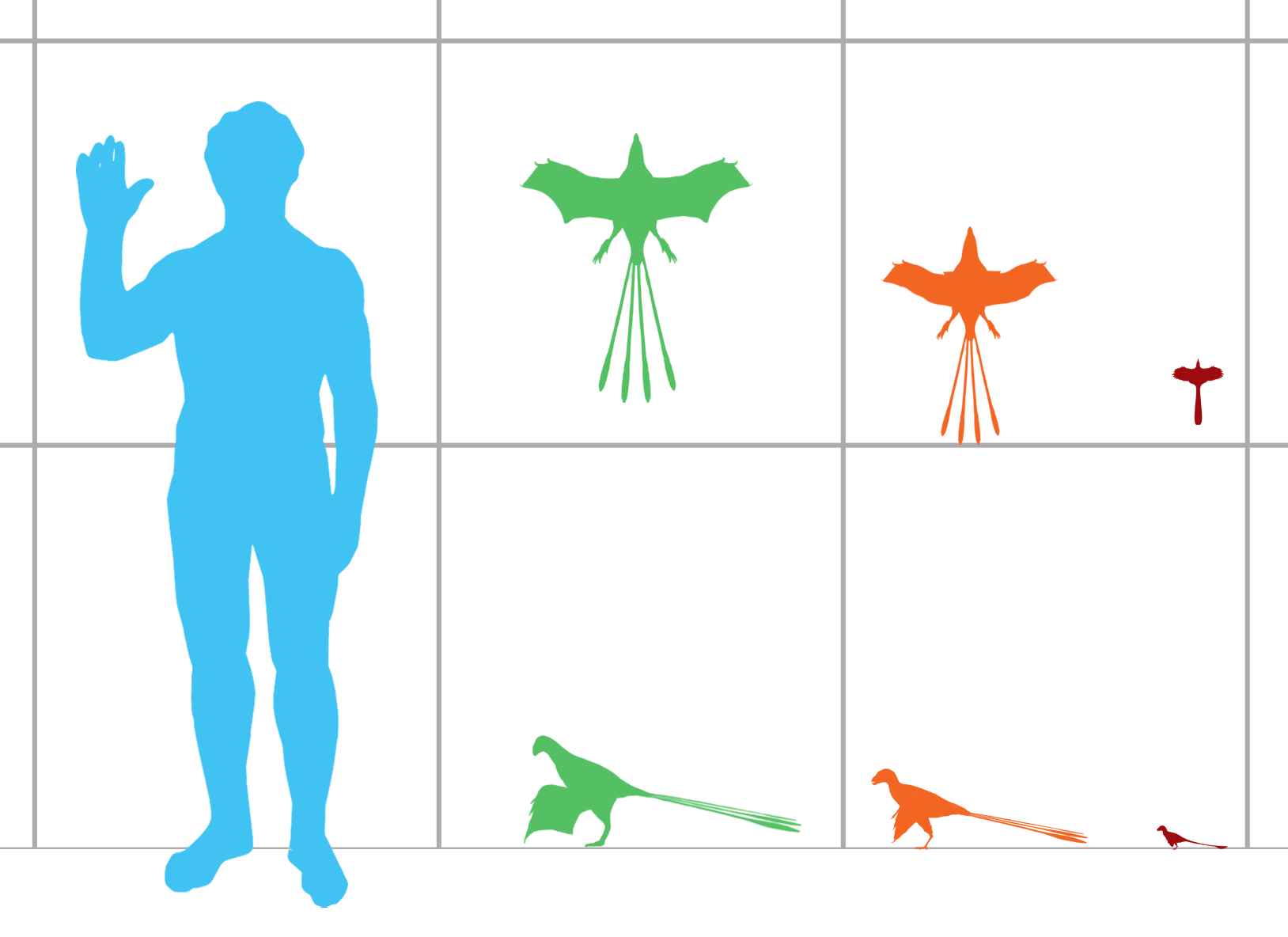
Other characteristics shared within the group include short, tall skulls with downturned lower jaws and large front teeth, and long arms. Tail length, however, varied significantly among scansoriopterygids. Epidexipteryx had a short tail (70% of torso length), anchoring long tail feathers, while Scansoriopteryx had a very long tail (more than three times as long as the torso) with a short spray of feathers at the tip. All three described scansoripterygid specimens retain trace fossil feathers covering their bodies. [2] [5] [6]

Scansoriopterygidae was created as a family -level taxon by Stephen Czerkas and Yuan Chongxi in 2002. Some scientists, such as Paul Sereno , initially considered the concept redundant because the group was originally monotypic, containing only the single genus and species Scansoriopteryx heilmanni . Furthermore, the group lacked a phylogenetic definition . [7] However, in 2008 Zhang et al. reported another scansoriopterygid, Epidexipteryx , and defined Scansoriopterygidae as a clade including the most recent common ancestor of Epidexipteryx and Epindrosaurus (=Scansoriopteryx ) plus all of its descendants. [6]

The exact taxonomic placement of this group was initially uncertain and controversial. Describing the first validly published specimen in 2002 ( Scansoriopteryx heilmanni ), Czerkas and Yuan hypothesized that various primitive skeletal features (including a primitive, “Saurischian-style” pubis and a primitive hip joint) show that scansoriopterygids, along with other maniraptorans and birds, separated from other theropods very early in dinosaur evolution. [5] However, this interpretation has not been followed by most other researchers. In a 2007 cladistic analysis of relationships among coelurosaurs , it was a member of the clade Avialae . [8] This view was supported by a second phylogenetic analysis performed by Zhang et al. in 2008.

A subsequent phylogenetic analysis by Agnolín and Novas (2011) recovered scansoriopterygids not as avialans, but as basal members of the clade Paraves remaining in unresolved polytomy with alvarezsaurids and the clade Eumaniraptora (containing avialans and deinonychosaurs ). [9]
Turner, Makovicky, and Norell (2012) only included Epidexipteryx hui in their primary phylogenetic analysis, as one adult specimen of this species is known; as for Scansoriopteryx / Epidendrosaurus , the authors were concerned that including it in the primary analysis would be problematic, because it is only known from juvenile specimens, which “do not necessarily retain all of the adult morphology needed to accurately place a taxon phylogenetically” (Turner, Makovicky and Norell 2012, p.89). Epidexipteryx was recovered as a basal paravia that did not belong to Eumaniraptora. The authors noted that its phylogenetic position is unstable; binding Epidexipteryx huias a basal avialan it required two additional steps compared to the more parsimonious solution, while constraining it as a basal member of Oviraptorosauria required only one additional step. A separate exploratory analysis included Scansoriopteryx / Epidendrosaurus , which was recovered as a basal member of Avialae; the authors noted that it has no clade with Epidexipteryx , which remained outside Eumaniraptora. Limiting the monophyly of Scansoriopterygidae required four additional steps and displaced Epidexipteryx into Avialae. [10]
A monophyletic Scansoriopterygidae was recovered by Godefroit et al. (2013); the authors found that scansoriopterygids were nearly basal members of Paraves and the sister group of the containing clade Avialae and Deinonychosauria. [11] Agnolín and Novas (2013) recovered scansoriopterygids as non-paravian maniraptorans and the sister group of Oviraptorosauria. [12] Brusatte et al. (2014) also found Epidexipteryx to be a basal oviraptorosaur along with Pedopenna . [13] Bayesian analysis by Cau (2018) re-placed scansoriopterygids in Oviraptorosauria, while parsimony analysis placed them in the basis of Avialae and included Xiaotingia in Scansoriopterygidae as a sister to the rest of the group. [14] Pitman et al . (2020) again found that scansoriopterygids were basal oviraptorosaurs. [15]
The following cladogram follows the results of a phylogenetic study by Lefèvre et al. , 2014: [16]
| Pennaraptor |
|
|||||||||||||||||||||||||||
Sorkin (2021) argues that scansoriopterygids provide evidence that all pennaraptorans evolved from scansorial gliding ancestors. [17]
Origin and palaeoecology
The fossil remains of Epidexipteryx , Scansoriopteryx , and Yi have all been recovered from the Tiaojishan Formation of northeastern China, and the first two have been found specifically in the Daohugou Beds . A study published in 2008 refined the possible age range of this formation, finding that the lower limit of the Tiaojishan was formed 165 Ma ago and the upper limit somewhere between 156 and 153 Ma ago. [18]
The known scansoriopterygids from the Daohugou biota inhabited a moist, temperate forest composed of a variety of prehistoric trees including ginkgo species and conifers . The understory would have been dominated by plants such as mosses , horsetails , cycads and ferns . [19]
Scansoriopterygids would have lived alongside synapsids such as the aquatic Castorocauda, Volaticotherium , a gliding arboreal mammal, and various types of gliding Haramiyidans , the rhamphorhynchoid pterosaurs Jeholopterus and Pterorhynchus , as well as a diverse range of insects (including mayflies and beetles ) and several species of salamander . [20] [21]
Paleobiology:
Climbing
In initial descriptions of the first two scansoriopterygid specimens, scientists studying these animals used several lines of evidence to argue that they were arboreal (tree climbers) and the earliest known non-avian dinosaurs with clear climbing adaptations.
Zhang and colleagues considered Scansoriopteryx arboreal based on the elongated nature of the hand and foot specializations. These authors argued that the long hand and strongly curved claws were adaptations for climbing and moving through tree branches. They saw it as an early stage in the evolution of the bird’s wing, stating that the forelimbs had become well developed for climbing and that this development later led to the evolution of a wing capable of flight. They argued that long, grasping hands are better suited for climbing than flying, as most flying birds have relatively short hands. Zhang et al. also noted that the foot of Scansoriopteryx it is unique among non-avian theropods; while Scansoriopteryx does not retain an inverted big toe (the rear-pointing toe seen in modern perching birds), its foot was very similar in construction to primitive perching birds such as Cathayornis and Longipteryx . These adaptations for grasping in all four limbs, the authors argued, make it likely that Scansoriopteryx spent a significant amount of time living in trees. [2]
In describing Scansoriopteryx , Czerkas and Yuan also described evidence for an arboreal lifestyle. They noted that, unlike all modern bird hatchlings, the forelimbs of Scansoriopteryx are longer than the hind limbs. The authors argued that this anomaly indicates that the forelimbs played an important role in locomotion even at an extremely early stage of development. Scansoriopteryx has a better preserved foot than the Epidendrosaurus type, and the authors interpreted the big toe as inverted, the condition of a backward-pointing toe being widespread among modern tree-dwelling birds. Furthermore, the authors pointed to the stiffened tail of Scansoriopteryx as a tree-climbing adaptation. The tail may have been used as a prop, much like the tails of modern woodpeckers . Comparison with the hands of modern climber species with an elongated third toe, such as iguanid lizards , also supports the tree-climbing hypothesis. In fact, Scansoriopteryx ‘s hands are much better adapted to climbing than the modern tree-climbing hoatzin pup. [5]
Feathers
Both juvenile scansoriopterygid specimens retain impressions of down-like simple feathers, especially around the hand and arm. The longer feathers in this region led Czerkas and Yuan to hypothesize that adult scansoriopterygids may have had reasonably well-developed wing feathers that could have aided in rudimentary jumping or gliding, although they ruled out the possibility that Scansoriopteryx could have achieved powered flight . Like other maniraptorans , scansoriopterygids had a carpal semilunate (crescent-shaped wrist bone) that allowed for a bird-like bending motion in the hand. Even if powered flight had not been possible, this movement could have aided maneuverability in jumping from branch to branch. [5]

Adult Epidexipteryx lacked preserved feathers around the forelimbs, but retained simple feathers on the body and long, ribbon-like feathers on the tail. The tail feathers, probably used in display, consisted of a central shaft (rachis) and an unbranched vane (unlike modern feather vanes, which are broken into smaller filaments or tips). [6]
Yi also keeps feathers. These are remarkably very simple for a member of Pennaraptora (a clade of which scansoriopterygids are usually considered members), being “brush-like”, with long, quill-shaped bases topped with sprays of finer filaments. Feathers covered most of the body, starting near the tip of the snout. The feathers on the head and neck were long and formed a thick coat, and the feathers on the body were even longer and denser, making it difficult for scientists to study their detailed structure.

Sliding membranes
At least two species, Yi qi and Ambopteryx longibrachium , have evolved a patagium , supporting it with an elongated third toe and a single styliform wrist bone resembling similar structures in flying squirrels , bats , pterosaurs , and anomalies . Although propatagi are known in birds and similar dinosaurs, scansoriopterygids were the only dinosaurs known to develop true membranous wings, particularly due to the presence of already quite derived feathers. [4] [22] [23] [24]
Prior to Yi ‘s discovery , Italian paleontologist Andrea Cau had informally suggested that membranes may be present in Scansoriopteryx , supported by its elongated third toe, due to their resemblance to the wing digits of pterosaurs, [25] a hypothesis that later he also applied to the Epidexipteryx. _ [26]

References
- ^ Hartman, Scott; Mortimer, Mickey Mouse; Wahl, William R.; Lomax, Dean R.; Lippincott, Jessica; Lovelace, David M. (2019). “A new paravian dinosaur from the Late Jurassic of North America supports a late takeover of avian flight”. PeerJ. 7:e7247. doi:10.7717/peerj.7247. PMC 6626525. PMID 31333906 .
- ^ Jump up to: a b c Zhang F.; Zhou Z.; Xu X.; Wang X. (2002). “A juvenile coelurosaur theropod from China indicates arboreal habits”. Naturwissenschaften . 89 (9):394–398. Bibcode : 2002NW…..89..394Z . doi : 10.1007/s00114-002-0353-8 . PMID 12435090 . S2CID 556221 .
- ^ Zhang, F.; Zhou, Z.; Xu, X.; Wang, X.; Sullivan, C. (2008). “A bizarre Jurassic maniraptoran from China with elongated ribbon-like feathers,” Supplementary Information.” Nature . 455 (7216): 1105–8. doi : 10.1038/nature07447 . PMID 18948955 . S2CID 4362560 .
- ^ Jump up to: a b Xu, X.; Zheng, X.; Sullivan, C.; Wang, X.; Xing, L.; Vanga, Y.; Zhang, X.; o’Connor, JK; Zhang, F.; Pan, Y. (2015). “A bizarre Jurassic maniraptoran theropod with preserved evidence of membranous wings”. Nature . 521 (7550): 70–3. Bibcode : 2015Natur.521…70X . doi : 10.1038/nature14423 . PMID 25924069 . S2CID 205243599 .
- ^ Jump up to: a b c d Czerkas, SA, and Yuan, C. (2002). “An arboreal maniraptoran from northeastern China.” pp. 63-95 in Czerkas, SJ (ed.), Feathered dinosaurs and the origin of flight. The Dinosaur Museum Journal 1 . The Dinosaur Museum, Blanding, USA Abridged version PDF
- ^ Jump up to: a b c Zhang, F.; Zhou, Z.; Xu, X.; Wang, X.; Sullivan, C. (2008). “A bizarre Jurassic maniraptoran from China with elongated ribbon-like feathers” (PDF) . Nature . 455 (7216): 1105–1108. doi : 10.1038/npre.2008.2326.1 . PMID 18948955 .
- ^ Clear, PC (2005). “ Scansoriopterygidae Archived 2008-01-27 at the Wayback Machine .” Stem Archosauria—TaxonSearch [version 1.0, November 7, 2005]
- ^ Senter, P (2007). “A new look at the phylogeny of Coelurosauria (Dinosauria: Theropoda)”. Journal of Systematic Paleontology . 5 (4):429–463. doi : 10.1017/S1477201907002143 . S2CID 83726237 .
- ^ Agnolin, Federico L.; Novas, Fernando E. (2011). “Unenlagiid theropods: are they members of the Dromaeosauridae (Theropoda, Maniraptora)?” . Anais from Academia Brasileira de Ciências . 83 (1):117–162. doi : 10.1590/S0001-37652011000100008 . PMID 21437379 .
- ^ Alan Hamilton Turner, Peter J. Makovicky and Mark Norell (2012). “A review of dromaeosaurid systematics and paravian phylogeny” . American Museum of Natural History Bulletin . 371 :1–206. doi : 10.1206/748.1 . hdl : 2246/6352 . S2CID 83572446 .
- ^ Pascal Godefroit , Helena Demuynck, Gareth Dyke, Dongyu Hu, François Escuillié and Philippe Claeys (2013). “Reduced Plumage and Flight Ability of a New Jurassic Paravian Theropod from China” . Communications Nature . 4 : Article number 1394. Bibcode : 2013NatCo…4.1394G . doi : 10.1038/ncomms2389 . PMID 23340434 .
- ^ Federico L. Agnolín and Fernando E. Novas (2013). Avian ancestors. A review of the phylogenetic relationships of the theropods Unenlagiidae, Microraptoria, Anchiornis and Scansoriopterygidae . SpringerBriefs in Earth System Science. pp. 1–96. doi : 10.1007/978-94-007-5637-3 . ISBN 978-94-007-5636-6. S2CID 199493087 .
- ^ Brusatte, Stephen L.; Lloyd, Graeme T.; Wang, Steve C.; Norell, Mark A. (2014-10-20). “Gradual assembly of avian body plan culminated in rapid rates of evolution during dinosaur-bird transition” . Current Biology . 24 (20): 2386–2392 . doi : 10.1016/j.cub.2014.08.034 . ISSN 1879-0445 . PMID 25264248 .
- ^ Cau, A. (2018). “The assembly of the avian body plan: a 160 million year process” . S2CID 44078918 .
- ^ Pitman, Michael; Xu, Xing (August 2020). “Pennaraptoran Theropod Dinosaurs Past Progress and New Frontiers” . American Museum of Natural History Bulletin . 440 (1):1–355. doi : 10.1206/0003-0090.440.1.1 . ISSN 0003-0090 .
- ^ Lefevre, U.; Hu, D.; Escuille, FO; Dam, G.; Godefroit, P. (2014). “A new basal long-tailed bird from the Early Cretaceous of northeastern China” . Biological Journal of the Linnean Society . 113 (3):790–804. doi : 10.1111/bij.12343 .
- ^ Sorkin, Boris (2021-12-02). “Scanning and Aerial Ability in Scansoriopterygidae and Basal Oviraptorosauria” . Historical Biology . 33 (12): 3202–3214 . doi : 10.1080/08912963.2020.1855158 . ISSN 0891-2963 . S2CID 230540120 .
- ^ Zhang, H.; Wang, M.; Liu, X. (2008). “Upper Boundary Age Constraints of Tiaojishan Formation Volcanic Rocks in Western Liaoning-North Hebei Using LA-ICP-MS Dating” . Chinese Science Bulletin . 53 (22): 3574–3584 . Bibcode : 2008SciBu..53.3574Z . doi : 10.1007/s11434-008-0287-4 .
- ^ Zhang K.; Yang D.; Dong R. (2006). “The Early Snipe (Diptera: Rhagionidae) from the Middle Jurassic of Inner Mongolia, China”. Zootaxa . 1134 :51–57. doi : 10.11646/zootaxa.1134.1.3 .
- ^ Meng J.; Hu Y.; Li C.; Wang Y. (2006). “The Mammal Fauna in the Early Cretaceous Jehol Biota: Implications for Mesozoic Mammal Diversity and Biology”. Geological Journal . 41 (3–4): 439–463. doi : 10.1002/gj.1054 . S2CID 84397126 .
- ^ Wang X.; Zhou Z. (2006). “Pterosaur assemblages from the Jehol Biota and their implication for Early Cretaceous pterosaur radiation”. Geological Journal . 41 (3–4): 405–418. doi : 10.1002/gj.1046 . S2CID 131602205 .
- ^ Yong, Ed (April 29, 2015). “Chinese dinosaur had bat-like wings and feathers” . National Geographic .
- ^ Greshko, Michael (8 May 2019). “New species of dinosaur with bat wings discovered” . National Geographic .
- ^ Minimum Wang; Jingmai K. O’Connor; XingXu; Zhonghe Zhou (2019). “A new Jurassic scansoriopterygid and the loss of membranous wings in theropod dinosaurs”. Nature. 569 (7755): 256–259. doi:10.1038/s41586-019-1137-z.
- ^ Cau, A (2008), Super Theropod Week, Part 1: Scansoriopterygid paleobiology, more than just bird origins! , Theropoda, October 2008
- ^ Cau, A (2012), Return of the paravian pterosaur-mime? , Theropoda, July 2012







