 Scansoriopterygidae (che significa “ali rampicanti”) è una famiglia estinta di dinosauri maniraptoran rampicanti e plananti . Gli scansoriopterigidi sono noti da cinque fossili ben conservati , che rappresentano quattro specie, rinvenuti nei letti fossili della Formazione Tiaojishan (risalenti al medio-tardo periodo Giurassico ) di Liaoning e Hebei , in Cina:
Scansoriopterygidae (che significa “ali rampicanti”) è una famiglia estinta di dinosauri maniraptoran rampicanti e plananti . Gli scansoriopterigidi sono noti da cinque fossili ben conservati , che rappresentano quattro specie, rinvenuti nei letti fossili della Formazione Tiaojishan (risalenti al medio-tardo periodo Giurassico ) di Liaoning e Hebei , in Cina:

Scansoriopteryx heilmanni (e il suo probabile sinonimo Epidendrosaurus ninchengensis ) è stato il primo dinosauro non aviario scoperto che aveva chiari adattamenti a uno stile di vita arboreo o semi-arboreo: è probabile che trascorressero gran parte del loro tempo sugli alberi. Entrambi gli esemplari mostravano caratteristiche che indicavano che erano giovani, il che rendeva difficile determinare la loro esatta relazione con altri dinosauri e uccelli non aviari . Non è stato fino alla descrizione di Epidexipteryx hui nel 2008 che era noto un esemplare adulto. Nel 2015 la scoperta di un altro esemplare adulto più grande appartenente alla specie Yi qi ha mostrato che gli scansoriopterigidi non erano solo scalatori, ma avevano anche adattamenti che avrebbero potuto essere usati per il volo planato. Lo dimostra anche l’Ambopteryx recentemente scoperto (nel 2019) . Anche la Pedopenna descritta in precedenza potrebbe essere un membro di questo clade. [1]

Gli scansoriopterigi sono tra i più piccoli dinosauri non aviari conosciuti. Gli esemplari giovanili di Scansoriopteryx hanno le dimensioni di passeri domestici , [2] lunghi circa 16 cm (6,3 pollici), mentre l’esemplare di tipo adulto di Epidexipteryx ha all’incirca le dimensioni di un piccione , lungo circa 25 cm (9,8 pollici) (escluso le penne della coda). [3]
Gli scansoriopterigidi si differenziano dagli altri dinosauri teropodi in parte per il loro terzo dito estremamente lungo, che era più lungo del primo e del secondo dito della mano. In tutti gli altri teropodi conosciuti, il secondo dito è il più lungo. Almeno due specie, Yi e Ambopteryx , avevano anche un lungo osso “stiliforme” che cresceva dal polso, che, insieme al terzo dito, aiutava a sostenere una membrana alare simile a un pipistrello usata per planare. Questo uso di un lungo dito per sostenere una membrana alare è solo superficialmente simile alla disposizione delle ali negli pterosauri , anche se fisicamente è più simile a un pipistrello. [4]
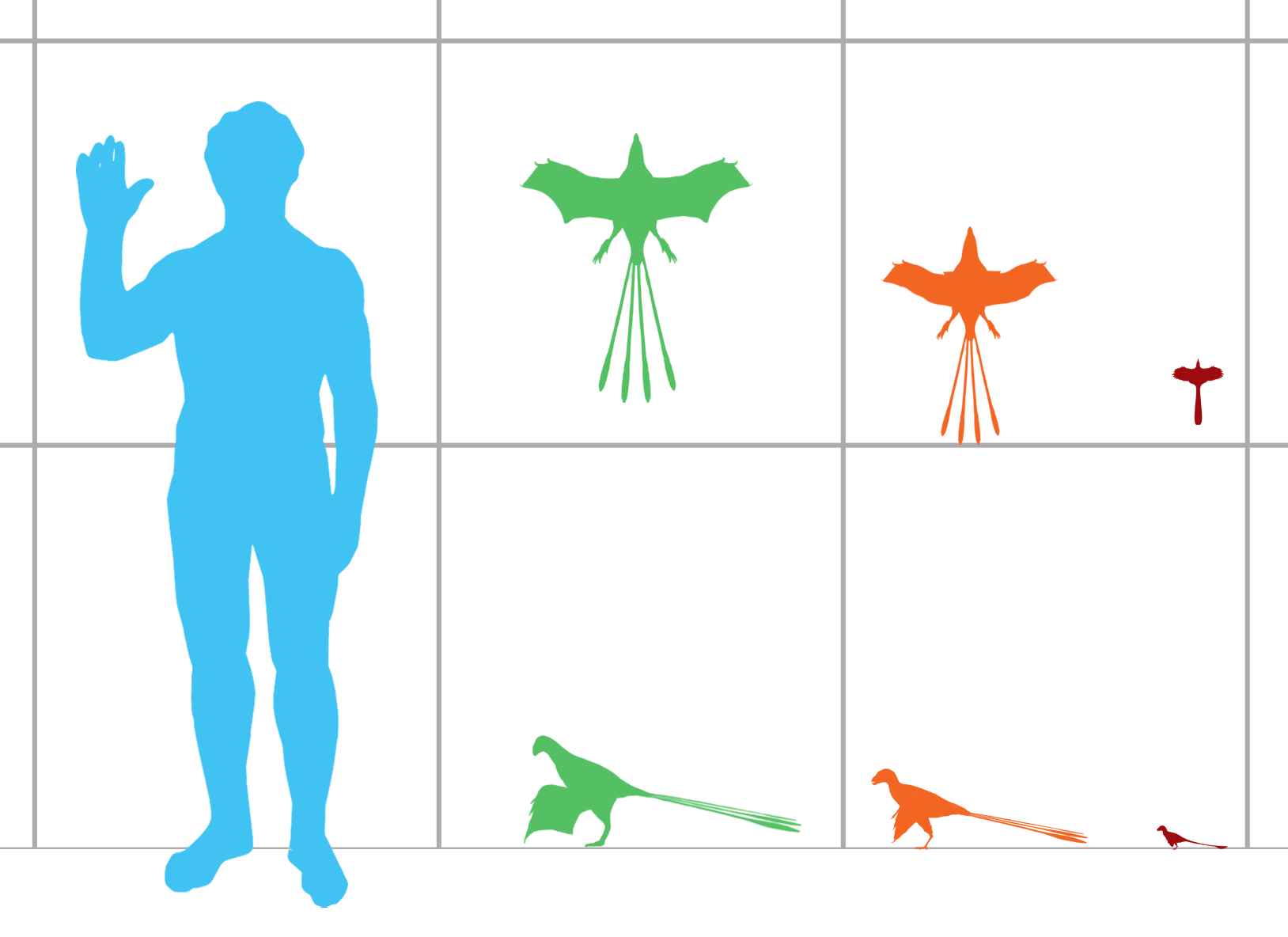
Altre caratteristiche condivise all’interno del gruppo includono teschi corti e alti con mascelle inferiori rivolte verso il basso e grandi denti anteriori e braccia lunghe. La lunghezza della coda, tuttavia, variava significativamente tra gli scansoriopterigidi. Epidexipteryx aveva una coda corta (il 70% della lunghezza del busto), ancorando lunghe penne della coda, mentre Scansoriopteryx aveva una coda molto lunga (più di tre volte più lunga del busto) con un breve spruzzo di piume sulla punta. Tutti e tre gli esemplari scansoripterigidi descritti conservano le tracce fossili di piume che ricoprono i loro corpi. [2] [5] [6]

Scansoriopterygidae è stato creato come taxon a livello familiare da Stephen Czerkas e Yuan Chongxi nel 2002. Alcuni scienziati, come Paul Sereno , inizialmente hanno considerato il concetto ridondante perché il gruppo era originariamente monotipico, contenente solo il singolo genere e la specie Scansoriopteryx heilmanni . Inoltre, il gruppo mancava di una definizione filogenetica . [7] Tuttavia, nel 2008 Zhang et al. riportato un altro scansoriopterygid, Epidexipteryx , e definito Scansoriopterygidae come un clade comprendente il più recente antenato comune di Epidexipteryx e Epidendrosaurus (=Scansoriopteryx ) più tutti i suoi discendenti. [6]

L’esatta collocazione tassonomica di questo gruppo era inizialmente incerta e controversa. Descrivendo il primo esemplare validamente pubblicato nel 2002 ( Scansoriopteryx heilmanni ), Czerkas e Yuan hanno ipotizzato che varie caratteristiche primitive dello scheletro (tra cui un pube primitivo, “in stile saurischiano” e un’articolazione dell’anca primitiva) mostrassero che gli scansoriopterigidi, insieme ad altri maniraptorani e uccelli, separati da altri teropodi molto presto nell’evoluzione dei dinosauri. [5] Tuttavia, questa interpretazione non è stata seguita dalla maggior parte degli altri ricercatori. In un’analisi cladistica del 2007 sulle relazioni tra i celurosauri , Phil Senter scoprì che Scansoriopteryx era un membro del clade Avialae .[8] Questo punto di vista è stato supportato da una seconda analisi filogenetica eseguita da Zhang et al. nel 2008.

Una successiva analisi filogenetica condotta da Agnolín e Novas (2011) ha recuperato scansoriopterygids non come avialans, ma come membri basali del clade Paraves rimanenti in politomia irrisolta con alvarezsauridi e il clade Eumaniraptora (contenente avialans e deinonychosaurs ). [9]
Turner, Makovicky e Norell (2012) hanno incluso solo Epidexipteryx hui nella loro analisi filogenetica primaria, poiché è noto un esemplare adulto di questa specie; per quanto riguarda Scansoriopteryx / Epidendrosaurus , gli autori erano preoccupati che includerlo nell’analisi primaria sarebbe stato problematico, perché è noto solo da esemplari giovani, che “non conservano necessariamente tutta la morfologia adulta necessaria per posizionare con precisione un taxon filogeneticamente” (Turner, Makovicky e Norell 2012, p.89). Epidexipteryx è stato recuperato come paravia basale che non apparteneva a Eumaniraptora. Gli autori hanno notato che la sua posizione filogenetica è instabile; vincolante Epidexipteryx huicome avialan basale richiedeva due passaggi aggiuntivi rispetto alla soluzione più parsimoniosa, mentre vincolarlo come membro basale di Oviraptorosauria richiedeva solo un passaggio aggiuntivo. Un’analisi esplorativa separata ha incluso Scansoriopteryx / Epidendrosaurus , che è stato recuperato come membro basale di Avialae; gli autori hanno notato che non ha clade con Epidexipteryx , che è rimasto al di fuori di Eumaniraptora. Limitare la monofilia di Scansoriopterygidae ha richiesto quattro passaggi aggiuntivi e ha spostato Epidexipteryx in Avialae. [10]
Uno Scansoriopterygidae monofiletico è stato recuperato da Godefroit et al. (2013); gli autori hanno scoperto che gli scansoriopterygidi erano quasi membri basali di Paraves e il gruppo gemello del clade contenente Avialae e Deinonychosauria. [11] Agnolín e Novas (2013) hanno recuperato gli scansoriopterigidi come maniraptorani non paraviani e il gruppo gemello di Oviraptorosauria. [12] Brusatte et al. (2014) hanno anche scoperto che Epidexipteryx è un oviraptorosauro basale insieme a Pedopenna . [13] L’analisi bayesiana di Cau (2018) ha collocato nuovamente gli scansoriopterygidi in Oviraptorosauria, mentre l’analisi della parsimonia li ha collocati nella base di Avialae e ha incluso Xiaotingiain Scansoriopterygidae come sorella del resto del gruppo. [14] Pitman et al . (2020) hanno nuovamente scoperto che gli scansoriopterigidi erano oviraptorosauri basali. [15]
Il cladogramma seguente segue i risultati di uno studio filogenetico di Lefèvre et al. , 2014: [16]
| Pennaraptor |
|
|||||||||||||||||||||||||||
Sorkin (2021) sostiene che gli scansoriopterigidi forniscono la prova che tutti i pennaraptorani si sono evoluti da antenati plananti scansoriali. [17]
Provenienza e paleoecologia
I resti fossili di Epidexipteryx , Scansoriopteryx e Yi sono stati tutti recuperati dalla formazione Tiaojishan della Cina nord-orientale, e i primi due sono stati trovati specificamente nei letti di Daohugou . Uno studio pubblicato nel 2008 ha perfezionato la possibile fascia di età di questa formazione, scoprendo che il limite inferiore del Tiaojishan si è formato 165 Ma fa e il limite superiore da qualche parte tra 156 e 153 Ma fa. [18]
I noti scansoriopterigidi del biota di Daohugou abitavano una foresta umida e temperata composta da una varietà di alberi preistorici tra cui specie di ginkgo e conifere . Il sottobosco sarebbe stato dominato da piante come muschi , equiseti , cicadee e felci . [19]
Gli scansoriopterygidi sarebbero vissuti accanto a sinapsidi come il Castorocauda acquatico, Volaticotherium , un mammifero arboricolo planante e vari tipi di haramiyidani plananti , gli pterosauri rhamphorhynchoid Jeholopterus e Pterorhynchus , oltre a una vasta gamma di insetti (tra cui effimere e coleotteri ) e diverse specie di salamandra . [20] [21]
Paleobiologia:
Arrampicata
Nelle descrizioni iniziali dei primi due esemplari scansoriopterygid, gli scienziati che studiano questi animali hanno utilizzato diverse linee di prova per sostenere che erano arboree (arrampicata sugli alberi) e i primi dinosauri non aviari conosciuti con chiari adattamenti all’arrampicata.
Zhang e colleghi consideravano Scansoriopteryx arboreo in base alla natura allungata della mano e alle specializzazioni del piede. Questi autori hanno affermato che la mano lunga e gli artigli fortemente ricurvi erano adattamenti per arrampicarsi e muoversi tra i rami degli alberi. Lo vedevano come una fase iniziale dell’evoluzione dell’ala dell’uccello, affermando che gli arti anteriori erano diventati ben sviluppati per l’arrampicata e che questo sviluppo in seguito portò all’evoluzione di un’ala capace di volare. Hanno sostenuto che le mani lunghe e prensili sono più adatte all’arrampicata che al volo, poiché la maggior parte degli uccelli volanti ha mani relativamente corte. Zhang et al. ha anche notato che il piede di Scansoriopteryx è unico tra i teropodi non aviari; mentre Scansoriopteryxnon conserva un alluce rovesciato (la punta rivolta all’indietro vista nei moderni uccelli appollaiati), il suo piede era molto simile nella costruzione ai primitivi uccelli appollaiati come Cathayornis e Longipteryx . Questi adattamenti per la capacità di afferrare in tutti e quattro gli arti, hanno sostenuto gli autori, rendono probabile che Scansoriopteryx abbia trascorso una notevole quantità di tempo a vivere sugli alberi. [2]
Nel descrivere Scansoriopteryx , Czerkas e Yuan hanno anche descritto prove di uno stile di vita arboricolo. Hanno notato che, a differenza di tutti i piccoli di uccelli moderni, gli arti anteriori di Scansoriopteryx sono più lunghi degli arti posteriori. Gli autori hanno sostenuto che questa anomalia indica che gli arti anteriori hanno svolto un ruolo importante nella locomozione anche in una fase di sviluppo estremamente precoce. Scansoriopteryx ha un piede meglio conservato rispetto al tipo di Epidendrosaurus e gli autori hanno interpretato l’alluce come invertito, la condizione di un dito rivolto all’indietro essendo diffusa tra i moderni uccelli che vivono sugli alberi. Inoltre, gli autori hanno indicato la coda irrigidita di Scansoriopteryxcome adattamento per arrampicarsi sugli alberi. La coda potrebbe essere stata usata come sostegno, proprio come le code dei picchi moderni . Anche il confronto con le mani delle moderne specie rampicanti con il terzo dito allungato, come le lucertole iguanidi , supporta l’ipotesi dell’arrampicata sugli alberi. In effetti, le mani di Scansoriopteryx sono molto più adatte all’arrampicata rispetto al moderno cucciolo di hoatzin che si arrampica sugli alberi . [5]
Piume
Entrambi gli esemplari giovanili di scansoriopterygid conservano impressioni di piume semplici simili a piumino, specialmente intorno alla mano e al braccio. Le piume più lunghe in questa regione hanno portato Czerkas e Yuan a ipotizzare che gli scansoriopterygidi adulti potessero avere penne delle ali ragionevolmente ben sviluppate che avrebbero potuto aiutare a saltare o planare rudimentali, sebbene escludessero la possibilità che Scansoriopteryx potesse aver raggiunto il volo potenziato . Come altri maniraptorani , gli scansoriopterigidi avevano un carpale semilunato (osso del polso a forma di mezzaluna) che consentiva un movimento di piegatura simile a un uccello nella mano. Anche se il volo a motore non fosse stato possibile, questo movimento avrebbe potuto aiutare la manovrabilità nel saltare da un ramo all’altro. [5]

L’esemplare adulto di Epidexipteryx mancava di piume conservate attorno agli arti anteriori, ma conservava piume semplici sul corpo e lunghe piume simili a nastri sulla coda. Le penne della coda, probabilmente utilizzate in esposizione, erano costituite da un fusto centrale (rachide) e da una paletta non ramificata (a differenza delle palette delle piume moderne, che sono spezzate in filamenti o punte più piccole). [6]
Yi conserva anche le piume. Questi sono notevolmente molto semplici per un membro di Pennaraptora (un clade di cui gli scansoriopterygids sono solitamente considerati membri), essendo “simili a un pennello”, con lunghe basi a forma di penna sormontate da spruzzi di filamenti più sottili. Le piume coprivano la maggior parte del corpo, iniziando vicino alla punta del muso. Le piume della testa e del collo erano lunghe e formavano uno spesso mantello, e le piume del corpo erano ancora più lunghe e dense, rendendo difficile per gli scienziati studiarne la struttura dettagliata.

Membrane scorrevoli
Almeno due specie, Yi qi e Ambopteryx longibrachium , hanno sviluppato un patagium , sostenendolo con il terzo dito allungato e un unico osso del polso stiliforme simile a strutture simili negli scoiattoli volanti , pipistrelli , pterosauri e anomalie . Sebbene i propatagi siano noti negli uccelli e in dinosauri simili, gli scansoriopterigidi erano gli unici dinosauri conosciuti a sviluppare vere ali membranose, in particolare a causa della presenza di piume già abbastanza derivate. [4] [22] [23] [24]
Prima della scoperta di Yi , il paleontologo italiano Andrea Cau aveva suggerito informalmente che le membrane potessero essere presenti in Scansoriopteryx , sostenute dal suo terzo dito allungato, a causa della loro somiglianza con le dita delle ali degli pterosauri, [25] un’ipotesi che in seguito applicò anche all’Epidexipterige . _ [26]

Riferimenti
- ^ Hartmann, Scott; Mortimer, Topolino; Wahl, William R.; Lomax, Dean R.; Lippincott, Jessica; Lovelace, David M. (2019). “Un nuovo dinosauro paraviano del tardo Giurassico del Nord America supporta un’acquisizione tardiva del volo aviario”. PeerJ. 7: e7247. doi:10.7717/peerj.7247. PMC 6626525. PMID 31333906 .
- ^Salta fino a:a b c Zhang F.; Zhou Z.; Xu X.; Wang X. (2002). “Un teropode celurosauro giovanile dalla Cina indica abitudini arboree”. Naturwissenschaften. 89(9): 394–398. Bibcode:2002NW…..89..394Z. doi:10.1007/s00114-002-0353-8. PMID12435090. S2CID556221.
- ^ Zhang, F.; Zhou, Z.; Xu, X.; Wang, X.; Sullivan, C. (2008). “Un bizzarro maniraptoran giurassico dalla Cina con piume allungate simili a nastri”, Informazioni supplementari “. Nature . 455 (7216): 1105–8. doi : 10.1038/nature07447 . PMID 18948955 . S2CID 4362560 .
- ^Salta fino a:a b Xu, X.; Zheng, X.; Sullivan, C.; Wang, X.; Xing, L.; Vanga, Y.; Zhang, X.; o’Connor, JK; Zhang, F.; Pan, Y. (2015). “Un bizzarro teropode maniraptoran giurassico con prove conservate di ali membranose”. Natura. 521(7550): 70–3. Bibcode:2015Natur.521…70X. doi:10.1038/nature14423. PMID25924069. S2CID205243599.
- ^Salta fino a:a b c d Czerkas, SA, e Yuan, C. (2002). “Un maniraptoran arboricolo del nord-est della Cina.” pp. 63-95 in Czerkas, SJ (a cura di),Dinosauri piumati e l’origine del volo. Il Diario del Museo dei Dinosauri1. The Dinosaur Museum, Blanding, USAVersione ridotta PDF
- ^Salta fino a:a b c Zhang, F.; Zhou, Z.; Xu, X.; Wang, X.; Sullivan, C. (2008). “Un bizzarro maniraptoran giurassico dalla Cina con piume allungate simili a nastri”(PDF). Natura. 455(7216): 1105–1108. doi: 10.1038/npre.2008.2326.1 . PMID18948955.
- ^ Sereno, PC (2005). “ Scansoriopterygidae Archiviato il 27-01-2008 su Wayback Machine .” Stem Archosauria—TaxonSearch [versione 1.0, 7 novembre 2005]
- ^ Senter, P (2007). “Un nuovo sguardo alla filogenesi di Coelurosauria (Dinosauria: Theropoda)”. Giornale di paleontologia sistematica . 5 (4): 429–463. doi : 10.1017/S1477201907002143 . S2CID 83726237 .
- ^ Agnolin, Federico L.; Novas, Fernando E. (2011). “Unenlagiid teropodi: sono membri dei Dromaeosauridae (Theropoda, Maniraptora)?” . Anais da Academia Brasileira de Ciências . 83 (1): 117–162. doi : 10.1590/S0001-37652011000100008 . PMID 21437379 .
- ^ Alan Hamilton Turner, Peter J. Makovicky e Mark Norell (2012). “Una revisione della sistematica dei dromaeosauridi e della filogenesi paraviana” . Bollettino del Museo Americano di Storia Naturale . 371 : 1–206. doi : 10.1206/748.1 . hdl : 2246/6352 . S2CID 83572446 .
- ^ Pascal Godefroit , Helena Demuynck, Gareth Dyke, Dongyu Hu, François Escuillié e Philippe Claeys (2013). “Piumeggio ridotto e capacità di volo di un nuovo teropode paraviano giurassico dalla Cina” . Comunicazioni Natura . 4 : Numero articolo 1394. Bibcode : 2013NatCo…4.1394G . doi : 10.1038/ncomms2389 . PMID 23340434 .
- ^ Federico L. Agnolín e Fernando E. Novas (2013). Antenati aviari. Una revisione delle relazioni filogenetiche dei teropodi Unenlagiidae, Microraptoria, Anchiornis e Scansoriopterygidae . SpringerBriefs in scienze del sistema terrestre. pp. 1–96. doi : 10.1007/978-94-007-5637-3 . ISBN 978-94-007-5636-6. S2CID 199493087 .
- ^ Brusatte, Stephen L.; Lloyd, Graeme T.; Wang, Steve C.; Norell, Mark A. (20-10-2014). “L’assemblaggio graduale del piano del corpo aviario è culminato in rapidi tassi di evoluzione durante la transizione dinosauro-uccello” . Biologia attuale . 24 (20): 2386–2392. doi : 10.1016/j.cub.2014.08.034 . ISSN 1879-0445 . PMID 25264248 .
- ^ Cau, A. (2018). “L’assemblaggio del piano del corpo aviario: un processo lungo 160 milioni di anni” . S2CID 44078918 .
- ^ Pitman, Michael; Xu, Xing (agosto 2020). “Pennaraptoran Theropod Dinosaurs Progresso passato e nuove frontiere” . Bollettino del Museo Americano di Storia Naturale . 440 (1): 1–355. doi : 10.1206/0003-0090.440.1.1 . ISSN 0003-0090 .
- ^ Lefèvre, U.; Hu, D.; Escuillé, FO; Diga, G.; Godefroit, P. (2014). “Un nuovo uccello basale dalla coda lunga del Cretaceo inferiore della Cina nord-orientale” . Giornale biologico della Linnean Society . 113 (3): 790–804. doi : 10.1111/bij.12343 .
- ^ Sorkin, Boris (2021-12-02). “Abilità scansoriale e aerea in Scansoriopterygidae e Oviraptorosauria basale” . Biologia Storica . 33 (12): 3202–3214. doi : 10.1080/08912963.2020.1855158 . ISSN 0891-2963 . S2CID 230540120 .
- ^ Zhang, H.; Wang, M.; Liu, X. (2008). “Vincoli sull’età del limite superiore delle rocce vulcaniche della formazione Tiaojishan nel Liaoning occidentale-Hebei settentrionale mediante datazione LA-ICP-MS” . Bollettino scientifico cinese . 53 (22): 3574–3584. Bibcode : 2008SciBu..53.3574Z . doi : 10.1007/s11434-008-0287-4 .
- ^ Zhang K.; Yang D.; Dong R. (2006). “Il primo beccaccino (Diptera: Rhagionidae) dal Giurassico medio della Mongolia Interna, Cina”. Zootaxa . 1134 : 51–57. doi : 10.11646/zootaxa.1134.1.3 .
- ^ Meng J.; Hu Y.; Li C.; Wang Y. (2006). “La fauna dei mammiferi nel Cretaceo inferiore Jehol Biota: implicazioni per la diversità e la biologia dei mammiferi mesozoici”. Giornale geologico . 41 (3–4): 439–463. doi : 10.1002/gj.1054 . S2CID 84397126 .
- ^ Wang X.; Zhou Z. (2006). “Assemblaggi di pterosauri del Jehol Biota e loro implicazione per la radiazione di pterosauri del Cretaceo inferiore”. Giornale geologico . 41 (3–4): 405–418. doi : 10.1002/gj.1046 . S2CID 131602205 .
- ^ Yong, Ed (29 aprile 2015). “Il dinosauro cinese aveva ali e piume simili a pipistrelli” . Geografico Nazionale .
- ^ Greshko, Michael (8 maggio 2019). “Scoperta nuova specie di dinosauro con ali di pipistrello” . Geografico Nazionale .
- ^ Minimo Wang; Jingmai K. O’Connor; XingXu; Zhonghe Zhou (2019). “Un nuovo scansoriopterygid giurassico e la perdita delle ali membranose nei dinosauri teropodi”. Natura. 569 (7755): 256–259. doi:10.1038/s41586-019-1137-z.
- ^ Cau, A (2008), Super Theropod Week, Parte 1: Scansoriopterygid paleobiology, più che semplici origini degli uccelli! , Theropoda, ottobre 2008
- ^ Cau, A (2012), ritorno del paraviano pterosauro-mimo? , Theropoda, luglio 2012







